Botany 2104 - Plant Form and Function
Respiration Overview
Cellular (Aerobic) Respiration
The overall reaction for respiration:
C6H12O6 + 6 O2 + 6 H2O ------------------------>
6 CO2 + 12 H2O
Basically, the process involves oxidizing sugar carbons to CO2. An
electron transport system is used to create a H+ (proton) gradient
that is used to make ATP. The final depository for the electrons is
O2, which is reduced to water.
Aerobic respiration is strongly identified with the mitochondria, however,
the starting reactions are in the cytosol. If oxygen is available,
the mitochondria get involved with respiration, and the internal structure
of the mitochondria is critical to the process.
Mitochondria
Two membranes: outer membrane and inner membrane
Small volume aqueous compartment: intermembrane space
Large volume aqueous compartment: matrix
The carbon oxidation takes place in two sets of reactions, glycolysis in
the cytosol and the Krebs cycle in the matrix of the mitochondria.
Both glycolysis and the Krebs cycle occur in steps. Stepwise oxidation
is important because:
1. it allows the dissipation of energy that is released as heat
2. it generates intermediates ==> steps to start making amino
acids, N-bases, other sugars for cell wall and nucleic acids, fatty acids,
chlorophyll, anthocyanins, hormones, secondary metabolites, etc.
Glycolysis
Glycolysis oxidizes glucose or fructose (C6 sugars) or triose-P (C3 sugars)
to pyruvate (C3 acid). Some of the energy released during the oxidation
can be captured as ATP. The electrons released by the oxidation are held
by NADH. Pyruvate is an organic acid. If the carbon is organic,
it is at least partly reduced. Therefore, there are still calories
(energy) available in pyruvate.
The Krebs cycle
The Krebs cycle finishes the carbon oxidation process. All of the carbons
in pyruvate are oxidized to CO2. It is a cyclic process, with the pyruvate
carbons getting attached to an existing cycle molecule prior to oxidation.
Once the pyruvate carbons are completely oxidized, the cycle molecule needs
to be regenerated. A little bit of ATP is made during the Krebs cycle,
but mostly it is an oxidation process. Most of the electrons are held
by NADH. A few are held by FADH2.
Electron Transport Chain (ETC)
The electrons from NADH and FADH2 are passed to a system of electron carriers
in the inner mitochondrial membrane, eventually reaching O2 which is then
reduced to H2O. As the electrons pass through the carriers, protons
(H+) are moved from the matrix to the intermembrane space. This establishes
a proton gradient. When the protons pass through a channel protein
back to the matrix, ATP is synthesized. How much ATP gets made?
It depends on who is doing the counting. Most plants that oxidize hexose molecule completely to CO2 will get between 30 and 38 ATP from glycolysis,
Krebs cycle, and ETC. The majority of that ATP (85-90%) comes from
the ETC. The high end of ATP yield (35-38 ATP) represents about
40% of the energy that was available in a molecule of glucose. The
remaining energy is given off as heat.
Some plants deliberately have an ATP yield in the mid-teens. Their
electrons take an alternative route in the ETC and don’t build as large as
proton gradient by the time they get to O2. Because the ATP yield is
down, these plants release more heat. These plants are called thermogenic.
They can raise the temperature in their microenvironment as much as 10oC.
This temperature increase volatilizes scents to attract pollinators more
efficiently.
Anaerobic respiration
Plants are able to respire anaerobically – without oxygen – for a short time.
Glycolysis still occurs, but any reactions involving the mitochondria are
not available. Without oxygen and the ETC, where can NADH unload its
electrons? Enter fermentation: a set of reactions that
let NADH unload electrons so glycolysis can continue. Most plants
use alcohol (ethanol) fermentation, but some use lactic acid fermentation,
and some use both types. The most likely anaerobic scenario for plants
==> roots in water-logged soil.
Some plants (hydrophytes) are specialized to have their roots always in water,
like anchored aquatic plants and emerged plants (water lilies, cattails,
etc.) The internal anatomy of these plants has large air spaces that
form passage ways to get O2 to the roots.
Respiration - Details
Respiration is essentially the cellular burning of a fuel to release its
energy. To burn the organic matter in a log, we would strike a match
to ignite the log and release the available energy as heat. In respiration,
the starting carbohydrate must be ignited; some of the released energy will
be as heat, but the rest of the energy will be captured as ATP. The
cellular equivalent of the match is ATP. The cell will have to spend
some existing ATP to start respiration, just as we need to use an existing
flame to ignite the log.
Glycolysis
The standard depiction of glycolysis is given in Figure 1. The process
starts with glucose. Two ATP are used to “ignite” each glucose molecule.
The end result is two molecules of triose-P, dihydroxyacetone phosphate (DHAP)
and phosphoglyceraldehyde (PGald). Energy has been expended, but no
energy has been collected. All of the carbon is present as carbohydrate,
which was the starting form of the carbon.
The next steps of glycolysis will oxidize the sugar carbons. The oxidation
process will release energy. Some of that energy will be captured as
4 ATP molecules. Some of the energy will be lost as heat. The
remainder of the released energy will still be potentially available.
That last portion of energy is associated with the electrons being held by
2 NADH molecules.
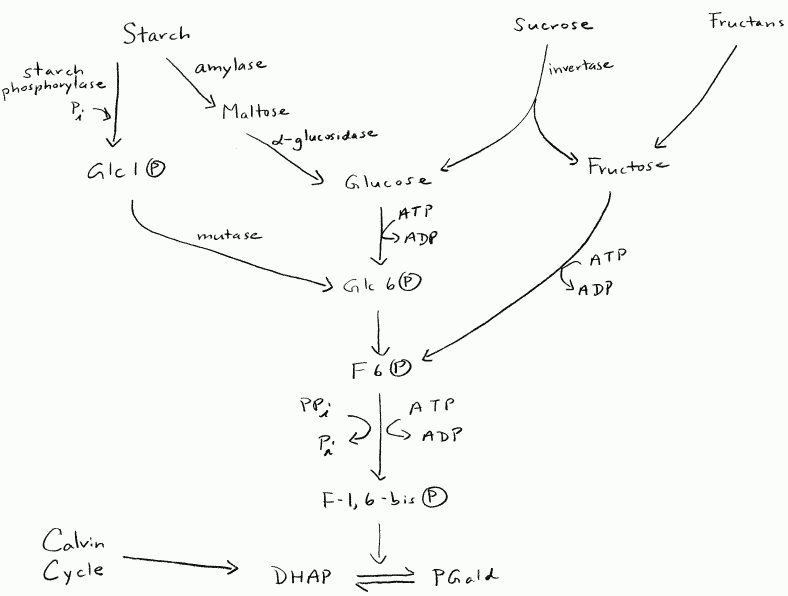
In general, plants don’t have a lot of free glucose lying around their cells.
Most of the sugars to be respired are in sucrose (arriving via the phloem),
newly made triose-P exported from the chloroplasts, and stored carbohydrate
polymers such as starch and fructans. Figure 2 shows how plants might
get carbohydrate from these sources to the triose-P step of glycolysis.
Plants don’t necessarily need to spend ATP to start the oxidation of sugars.
Glucose can be phosphorylated as it is released from starch by the action
of starch phosphorylase. There is an alternate enzyme for converting
fructose-6-P to fructose-1,6-bisP that uses inorganic pyrophosphate instead
of ATP.
Also, plants don’t necessarily make ATP during the oxidation steps of glycolysis.
It all depends on their phosphate status.
The reality of ATP yield for plants from glycolysis of one molecule of glucose
equivalent: -2 to + 4.
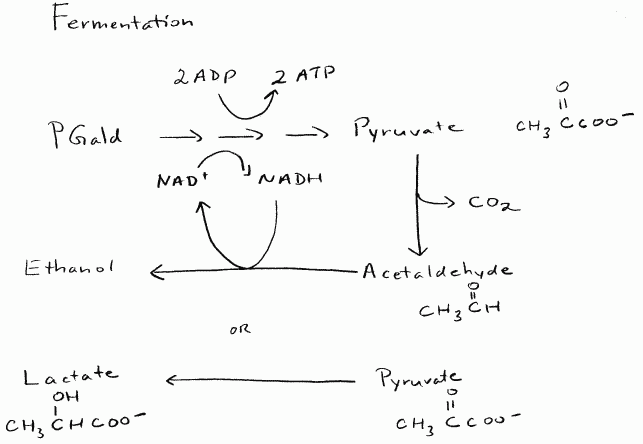
If oxygen is not present, the pyruvate that remains at the end of glycolysis
will be fermented. Fermentation allows the cell to unload the electrons
that were given to NAD+ during the oxidation steps of glycolysis. This
will free up the NAD+ molecules to get more electrons and keeps glycolysis
going. As fermentation is essentially a carbon reduction step, no additional
energy is released or captured as ATP in the process. The possibilities
for fermentation in plant cells are shown in Figure 4.
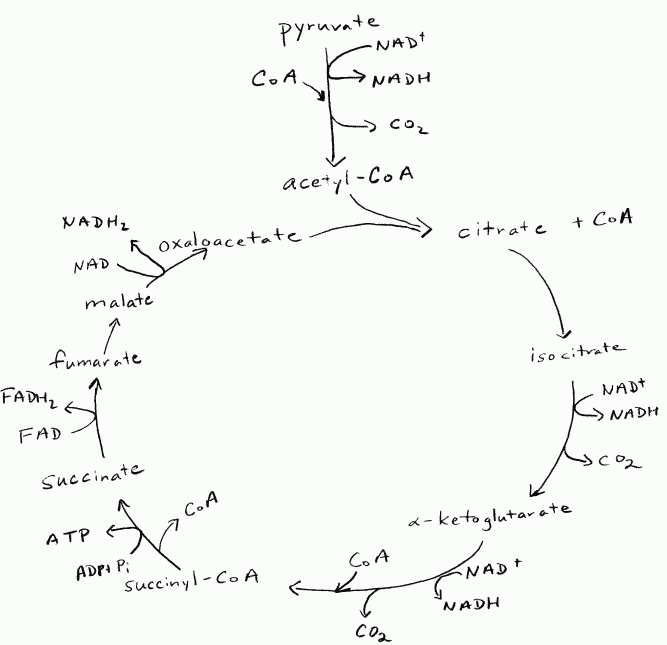
If oxygen is present, the oxidation of the carbons in pyruvate to CO2
continues in the matrix of the mitochondrion. This is where the Krebs
cycle (TCA cycle, citric acid cycle; Figure 5) is
located. Pyruvate is initially oxidized to release one carbon as CO2
with a C2 acetyl group to be attached to coenzyme A (CoA). The
oxidation step also results in the reduction of NAD+ to NADH.
The acetyl group is joined with an existing molecule of oxaloacetate (C4) to
give citrate (C6). Citrate is oxidized to α-ketoglutarate (C5), with the
reduction of NAD+ to NADH and the release of one carbon as CO2.
α-Ketoglutarate is converted to succinate (C4), with the release of one carbon
as CO2 and the phosphorylation of one ADP to ATP. Succinate is
oxidized to fumarate (C4), with the reduction of FAD to FADH2.
Following the conversion of fumarate to malate (C4), the malate is oxidized to
oxaloacetate (C4), with the reduction of NAD+ to NADH The
cycle is now reset to take in an acetyl group from the next pyruvate.
Summary of the Krebs cycle:
pyruvate + ADP + Pi + 4 NAD+ + FAD --------> 3 CO2
+ ATP + 4 NADH + FADH2
The Krebs cycle happens twice per glucose equivalent that started glycolysis.
Krebs cycle + glycolysis:
1. hexose ------> 6 CO2
2. phosphorylation of 0-6 ADP to 0-6 ATP
3. The oxidation of hexose released 24 electrons, now being held by 2 NADHG,
8 NADHK, and 2 FADH2. (The G and K subscripts on
NADH indicate that these molecules were reduced during glycolysis and the Krebs
cycle, respectively.)
Notice that aerobic respiration, which required molecular oxygen, hasn’t used
any oxygen yet. The ETC (Figure 6) will
transport the electrons from the dinucleotides to O2. The O2
will be reduced to water.
6 O2 + 24 electrons + 24 H+
--------> 12 H2O
As the electrons move through the ETC from the dinucleotides to O2, a
proton gradient is established. A high concentration of protons builds up
in the intermembrane space. These protons will diffuse through an ATPase
located in the inner membrane of the mitochondrion. As the protons pass
through the ATPase, its configuration changes. The enzyme is then able to
attach a free phosphate to an ADP, forming ATP.
The steeper the proton gradient, the more ATP that can be made.
Figure 6 shows that protons can pass from the
mitochondrial matrix to the intermembrane space at three points in the ETC.
Only electrons that are obtained from NADHK and given to Complex I
will travel through all three proton passage points. Electrons from NADHG
and FADH2 only travel through two proton passage points.
Various calculations and measurements show that a maximum of three ATP are made
per NADHK and two are made per NADHG or FADH2.
The actual number of ATP made depends on many factors, such as cellular demands
for reducing power elsewhere, whether NADHK gives its electrons
to Complex I or NADH dehydrogenase (B in Figure 6), and whether or not the
alternative oxidase is used.
Inhibitors of the ETC:
Complex I: rotenone, piericidin
Complex II: TTFA (thenoyltrifluoroacetone)
(succinate —> fumarate: malonate)
Complex III: antimycin
Complex IV: CO, CN-, N3-
Uncouplers: collapse the proton gradient. Oxygen is still reduced to
water, but no ATP can be made. dinitrophenol (DNP)
Figure 7 is an abbreviated version of glycolysis and
the Krebs cycle. These two pathways generate many intermediate molecules
as hexose is oxidized to CO2. At any point in the pathways, the
intermediates can be removed to make other molecules that the plant needs,
including both primary and secondary metabolites.
Review
Return to Botany 2104 Home Page.
Return to Botany 2104 Schedule.
27 July 2011
![]()
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}